| Issue |
Wuhan Univ. J. Nat. Sci.
Volume 31, Number 2, April 2026
|
|
|---|---|---|
| Page(s) | 101 - 111 | |
| DOI | https://doi.org/10.1051/wujns/2026312101 | |
| Published online | 13 May 2026 | |
Aquatic Ecology and Water Environment Safety
CLC number: O175.25
Qualitative Analysis and Numerical Simulation of a Predator-Prey Model for Invertebrate Predators in Aquatic Population Ecosystem
水生种群生态系统中具有无脊椎捕食者的捕食食饵模型定性分析与数值模拟
School of Mathematics and Information Science, Baoji University of Arts and Sciences, Baoji 721013, Shaanxi, China
(宝鸡文理学院 数学与信息科学学院, 陕西 宝鸡721013)
† Corresponding author. E-mail: This email address is being protected from spambots. You need JavaScript enabled to view it.
Received:
20
April
2025
Abstract
The predation mechanism of invertebrates (e.g., Tortanus dextrilobatus) on plankton in aquatic population ecosystem is a significant research topic. In this paper, the interaction between invertebrates and plankton is simulated by a modified Leslie-Gower predator-prey model. Using the theory of reaction-diffusion equations, a priori estimate, existence, uniqueness and stability conditions of the positive steady state solution are established. Furthermore, numerical simulations are conducted to quantitatively analyze the dynamical behavior. The research shows that as long as the Allee effect constant satisfies the appropriate relationship and the growth rates of predator and prey are appropriately large, the predator and prey can not only coexist, but also the coexistence mode is unique and stable under low predation-rate. In addition, the numerical simulations show that the coexistence may be stable under high predation-rate. Meanwhile, with the increase of predation rate, the population density of predators will decrease.
摘要
无脊椎动物(例如溞溏水蚤)对水生种群生态系统中浮游生物的捕食机制是一个重要的研究课题。本文采用改进的Leslie-Gower捕食-食饵模型模拟了无脊椎动物和浮游生物之间的相互作用。利用反应扩散方程理论,建立了正稳态解的先验估计、存在性、惟一性和稳定性条件。此外,利用数值模拟定量分析了模型动力学行为。研究表明,只要Allee效应常数满足适当的关系、捕食者和食饵的生长速度适当大,捕食者和食饵不仅可以共存,而且在低捕食率下共存模式是惟一且稳定的。此外,数值模拟表明,在高捕食率下,也可能共存。同时,随着捕食率的增加,捕食者的种群密度会减小。
Key words: predator-prey model / aquatic population ecosystem / Allee effect / Ivlev functional response / positive steady-state solutions / numerical simulation
关键字 : 捕食-食饵模型 / 水生种群生态系统 / Allee效应 / Ivlev型功能反应函数 / 正稳态解 / 数值模拟
Cite this article:WANG Lijuan, JIANG Hongling, HE Jinchan, et al. Qualitative Analysis and Numerical Simulation of a Predator-Prey Model for Invertebrate Predators in Aquatic Population Ecosystem[J]. Wuhan Univ J of Nat Sci, 2026, 31(2): 101-111.
Biography: WANG Lijuan, female, Master candidate, research direction: partial differential equations. E-mail:This email address is being protected from spambots. You need JavaScript enabled to view it.
Foundation item: Supported by the National Natural Science Foundation of China (11961030) , the Natural Science Foundation of Shaanxi Province (2022JM-034)
© Wuhan University 2026
 This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
0 Introduction
Let's start with the following predator-prey model
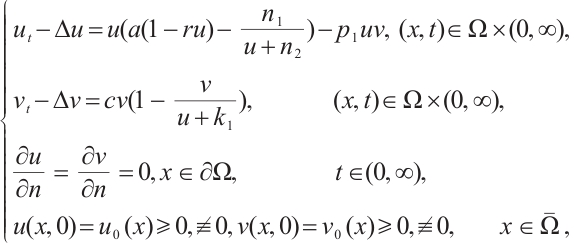 (1)
(1)
where 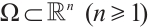 is a bounded domain with smooth boundary
is a bounded domain with smooth boundary  . In Ref. [1], the authors studied the predator-prey model (1) and gave the conditions for the steady-state bifurcation and Hopf bifurcation from the unique positive constant solution. The
. In Ref. [1], the authors studied the predator-prey model (1) and gave the conditions for the steady-state bifurcation and Hopf bifurcation from the unique positive constant solution. The 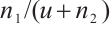 used here named additive Allee effect, where
used here named additive Allee effect, where 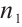 and
and 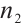 are constants of Allee effect which describing the strength of Allee effect, refers to that the low density populations may have difficulties in finding mates, promoting reproduction, predation, environmental regulation and inbreeding, which may lead to population extinction. Therefore, it is significant to study the mechanism of Allee effect to avoid population extinction in low density population. Some classic studies on the impact of Allee effect on predation models can be found in Refs. [2-5]. The background and parameter meanings of this model can be found in Ref. [1] and we omit here.
are constants of Allee effect which describing the strength of Allee effect, refers to that the low density populations may have difficulties in finding mates, promoting reproduction, predation, environmental regulation and inbreeding, which may lead to population extinction. Therefore, it is significant to study the mechanism of Allee effect to avoid population extinction in low density population. Some classic studies on the impact of Allee effect on predation models can be found in Refs. [2-5]. The background and parameter meanings of this model can be found in Ref. [1] and we omit here.
It is well known that the predator-prey functional response is an important factor in predator-prey models, which profoundly affects the dynamics of predator-prey models[6-7]. The functional response 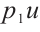 in (1) is linear function and is unbounded. This deficiency reminds us that we can use the following Ivlev function, which can be written as
in (1) is linear function and is unbounded. This deficiency reminds us that we can use the following Ivlev function, which can be written as 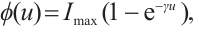 where
where 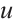 is the population density of prey,
is the population density of prey, 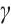 and
and 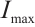 are positive constants which represent the predation rate of the predator and the maximum capture rate, respectively. It is clear that the
are positive constants which represent the predation rate of the predator and the maximum capture rate, respectively. It is clear that the 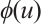 is bounded and satisfies
is bounded and satisfies 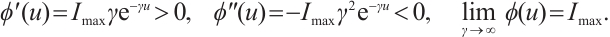
A recent experiment shows that the predation rate of Tortanus dextrilobatus expressed by 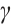 can simulate the model that Tortanus dextrilobatus prey on zooplankton in the San Francisco Estuary[8]. Many studies, both modeling analysis and ecological experiments, such as Refs. [8-11], show that the predation rate
can simulate the model that Tortanus dextrilobatus prey on zooplankton in the San Francisco Estuary[8]. Many studies, both modeling analysis and ecological experiments, such as Refs. [8-11], show that the predation rate  strongly affects the coexistence of predator and prey. However, there has been no research on using the Ivlev functional response to simulate predation of zooplankton in (1). For the sake of simplicity, we will take
strongly affects the coexistence of predator and prey. However, there has been no research on using the Ivlev functional response to simulate predation of zooplankton in (1). For the sake of simplicity, we will take 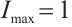 in our work. By introducing the following non-dimensional variables
in our work. By introducing the following non-dimensional variables
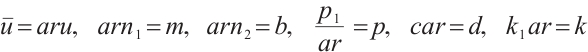
in (1) and dropping the superscripts of 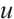 for simplicity, the predator-prey model (1) is given by the following equations
for simplicity, the predator-prey model (1) is given by the following equations
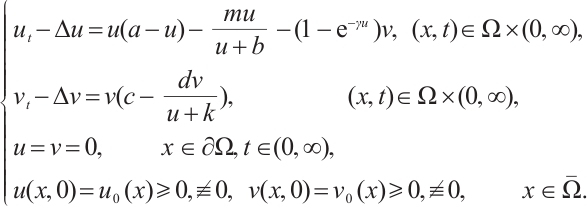 (2)
(2)
Here 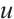 and
and 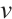 are the population densities of prey and predator,
are the population densities of prey and predator, 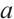 and
and 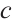 are the intrinsic growth rates of prey and predator, respectively. The Allee effect constants
are the intrinsic growth rates of prey and predator, respectively. The Allee effect constants 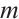 and
and 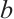 satisfy
satisfy 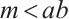 which is called weak additive Allee effect.
which is called weak additive Allee effect. 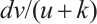 is a modified Leslie-Gower term[12-13]. The parameter
is a modified Leslie-Gower term[12-13]. The parameter 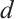 represents the maximum average reduction rate obtained by predators, and
represents the maximum average reduction rate obtained by predators, and 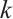 represents the environmental carrying capacity of predators. For more detailed biological significance of the model, one can see Refs. [14-16]. Obviously, the corresponding steady-state problem to (2) can be written as
represents the environmental carrying capacity of predators. For more detailed biological significance of the model, one can see Refs. [14-16]. Obviously, the corresponding steady-state problem to (2) can be written as
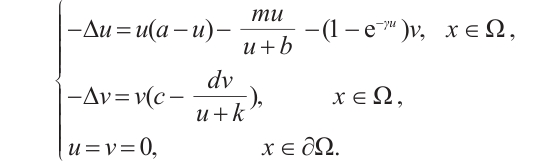 (3)
(3)
The main purpose of this paper is to clarify the influence of weak additive Allee effect and Ivlev functional response on the positive solution of model (2). The content of this paper is arranged as follows. Section 1 introduces some preliminary results. Section 2 gives the necessary conditions and prior estimates for positive solutions of (3). Section 3 gives the sufficient conditions for the existence of positive solutions of (3). Section 4 gives the uniqueness and stability of the positive solution of (3). In Section 5, the dynamics of (2) and (3) are quantitatively analyzed by numerical simulations.
1 Preliminaries
Let 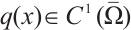 . It is well known that the following problem
. It is well known that the following problem
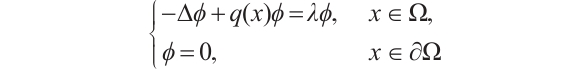 (4)
(4)
has an infinite sequence of eigenvalues which are bounded below. Throughout this paper, we denote the first eigenvalue by 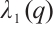 and the corresponding eigenfunction does not change sign on
and the corresponding eigenfunction does not change sign on 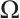 . We also denote that
. We also denote that 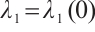 with the corresponding eigenfunction
with the corresponding eigenfunction 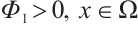 . For more detailed information, one can see Refs. [17-19].
. For more detailed information, one can see Refs. [17-19].
Now we consider the following boundary value problem
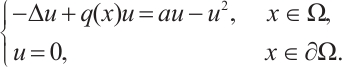
According to Ref. [20], if 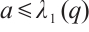 , then u=0 is the unique non-negative solution of this problem, and it has a unique positive solution if
, then u=0 is the unique non-negative solution of this problem, and it has a unique positive solution if 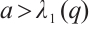 . In particular, if
. In particular, if 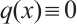 and
and 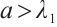 , then it has a unique positive solution, denoted by
, then it has a unique positive solution, denoted by 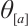 , which is monotonically increasing with respect to
, which is monotonically increasing with respect to 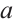 . And then, for the boundary value problem
. And then, for the boundary value problem
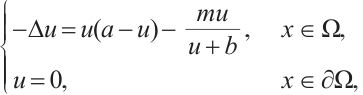 (5)
(5)
using the upper and lower solution method, it has a unique positive solution 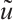 which satisfies
which satisfies 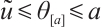 when
when 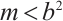 and
and 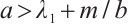 . We remark here that the condition
. We remark here that the condition 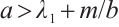 can meet the requirements of the condition of weak additive Allee effect, i.e.,
can meet the requirements of the condition of weak additive Allee effect, i.e.,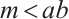 . Finally, consider the following boundary value problem
. Finally, consider the following boundary value problem
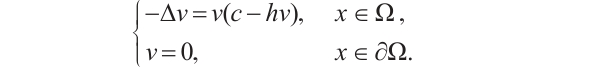 (6)
(6)
It has a unique positive solution, denoted by 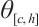 , if
, if 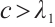 . Moreover,
. Moreover, 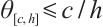 and is monotonically decreasing with respect to
and is monotonically decreasing with respect to 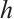 . Especially, when
. Especially, when 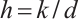 , denote the unique positive solution by
, denote the unique positive solution by 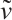 , that is
, that is 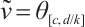 .
.
According to (5) and (6), (3) has a unique semi-trivial solution 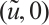 if
if 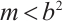 and
and 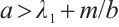 , and has a unique semi-trivial solution
, and has a unique semi-trivial solution 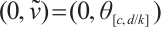 and
and 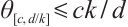 if
if 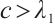 .
.
Lemma 1[21-22]
Let 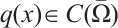 ,
,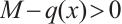 with the constant
with the constant 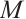 ,
, 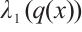 be the principal eigenvalue of (4). We have the following statements:
be the principal eigenvalue of (4). We have the following statements:
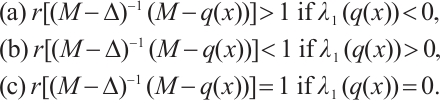
Lemma 2[22] Let 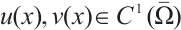 satisfy
satisfy
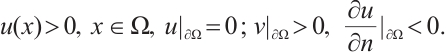
Then there exists 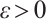 such that
such that 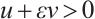 for any
for any  .
.
2 Necessary Conditions and Prior Estimates
In this section, we use the upper and lower solution method and the strong maximum principle to establish the necessary condition and a priori estimate of positive solutions of (3).
Theorem 1 If (3) has a positive solution, then 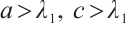 .
.
Proof Let 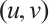 be a positive solution of (3). Multiply both sides of the first equation of (3) by
be a positive solution of (3). Multiply both sides of the first equation of (3) by 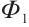 and integrate on
and integrate on 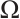 , we can get
, we can get
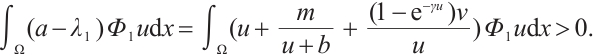
This implies that 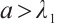 . The inequality
. The inequality 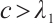 can be obtained by the second equation of (3) similarly.
can be obtained by the second equation of (3) similarly.
Remark 1 Theorem 1 shows that when the growth rate of predator or prey is low, at least one species is extinct in (3).
Theorem 2 Suppose that 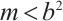 ,
,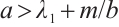 and
and 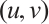 is a positive solution of (3). Then
is a positive solution of (3). Then
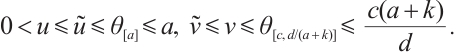
Proof The above two inequalities are proved in the same way, and we only prove the second inequality. According to the second equation of (3) we have
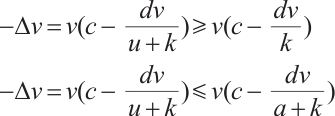
If 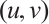 is a positive solution of (3), according to Theorem 1 we have
is a positive solution of (3), according to Theorem 1 we have 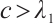 . Then
. Then
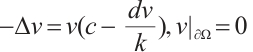 and
and 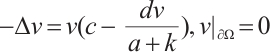
have a unique positive solution 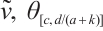 , respectively. Thus
, respectively. Thus 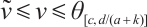 can be obtained from upper-lower solution method and uniqueness of
can be obtained from upper-lower solution method and uniqueness of 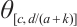 . The inequality
. The inequality 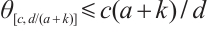 can be obtained from the nature of
can be obtained from the nature of 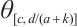 .
.
3 Existence of Positive Steady-State Solutions
In this section, we establish the existence of positive solution of (3) by using the degree theory. In order to apply the degree theory, we make the following definitions:
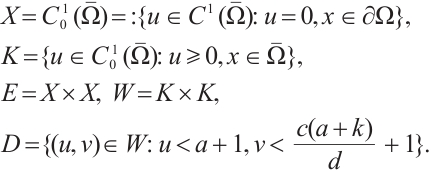
Using Lemma 2, we can get
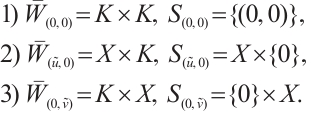
Lemma 3[22] For the mapping  . Suppose that
. Suppose that 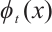 is continuous on
is continuous on 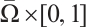 and
and 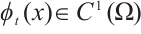 for any
for any 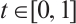 . If
. If 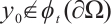 for any
for any 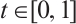 , then the topological degree
, then the topological degree 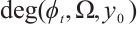 does not depend on
does not depend on 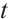 .
.
Lemma 4[22] Let 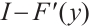 be invertible on
be invertible on 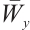 .
.
(i) If 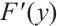 has
has 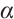 -property, then
-property, then 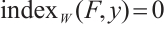 ;
;
(ii) If 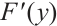 has no
has no 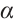 -property, then
-property, then 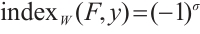 , where
, where 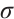 is the sum of the algebraic multiplicities of the eigenvalues of
is the sum of the algebraic multiplicities of the eigenvalues of 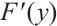 which are larger than one.
which are larger than one.
Lemma 5 Let 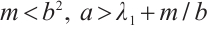 . Then all eigenvalues of
. Then all eigenvalues of 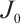 are greater than 0. Here
are greater than 0. Here 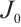 is the linearization operator of (5) at
is the linearization operator of (5) at 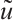 , i.e.,
, i.e.,
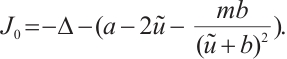
Proof By 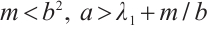 , we have
, we have 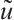 is a unique positive solution of (5). Thus
is a unique positive solution of (5). Thus
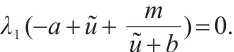
According to 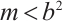 , we have
, we have
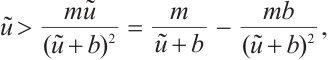
It means that 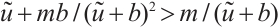 . By the nature of principle eigenvalue, there holds
. By the nature of principle eigenvalue, there holds
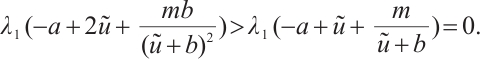
The proof is completed.
According to Theorem 2, we have any nonnegative solution of (3) belongs to  . Then there exists a positive constant
. Then there exists a positive constant  , such that
, such that
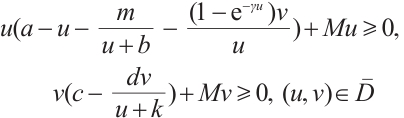
Define mapping 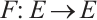 as
as
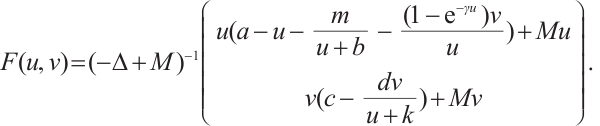 (7)
(7)
Then it is a compact operator and 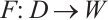 . Thus
. Thus 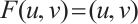 if and only if
if and only if 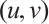 is a solution of (3).
is a solution of (3).
For any 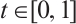 , we also define
, we also define
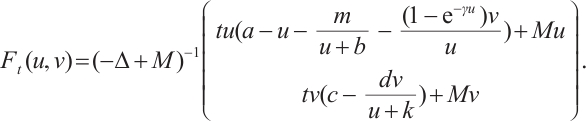
Clearly, 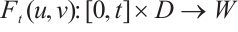 is a positive compact operator and
is a positive compact operator and 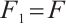 .
.
Lemma 6 Let 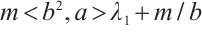 . We have the following statements:
. We have the following statements:
(i) 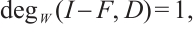
(ii) If 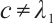 , then
, then 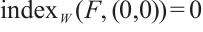 ,
,
(iii) If 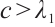 , then
, then 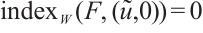 ,
,
(iv) If 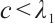 , then
, then 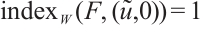 .
.
Proof (i) It is clear that 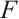 has no fixed point on
has no fixed point on  . For any
. For any 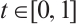 , the fixed point of
, the fixed point of 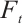 is equivalent to the solution of boundary value problem
is equivalent to the solution of boundary value problem
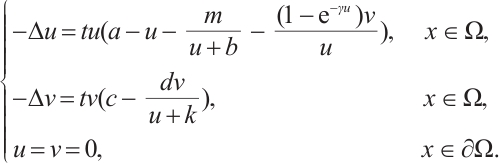
According to Theorem 2, the fixed point of 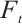 satisfies
satisfies 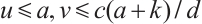 for any
for any 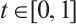 . Thus
. Thus 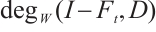 does not depend on
does not depend on 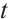 from homotopic invariant property and
from homotopic invariant property and
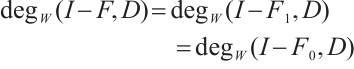
By the above boundary value problem has a unique solution 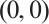 as
as 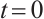 , we have
, we have
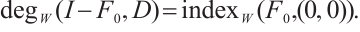
Notice 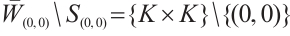 . Denote
. Denote 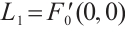 . Then
. Then
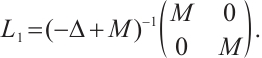
By 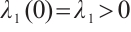 and Lemma 1,
and Lemma 1, 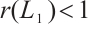 . It can derive that
. It can derive that 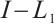 is invertible on
is invertible on 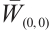 and
and 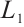 has no
has no 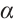 -property on
-property on 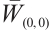 . According to Lemma 4, we have
. According to Lemma 4, we have
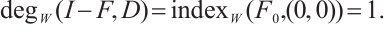
(ii) Let 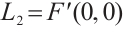 . Then
. Then
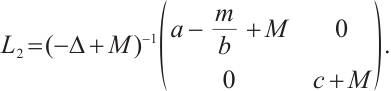
At first, we will show that 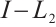 is invertible on
is invertible on 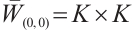 . If it is not true, then there is
. If it is not true, then there is 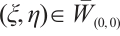 and
and 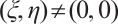 such that
such that 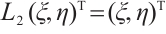 , i.e.,
, i.e.,
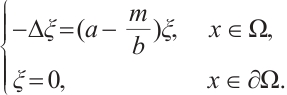
If 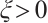 , then
, then 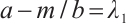 , this is contrary to
, this is contrary to 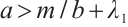 . Thus
. Thus  . Similarly,
. Similarly,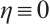 . This is a contradiction with
. This is a contradiction with 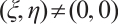 . Then
. Then 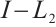 is invertible on
is invertible on 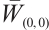 .
.
Now we claim that 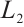 has
has 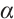 -property on
-property on 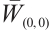 . By
. By 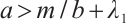 and Lemma 1,
and Lemma 1,
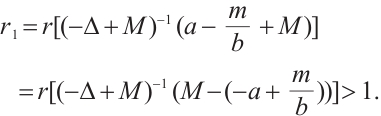
Notice that 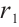 is the principal eigenvalue of the operator
is the principal eigenvalue of the operator 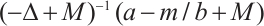 , and the corresponding eigenfunction is
, and the corresponding eigenfunction is 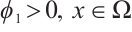 . Take
. Take 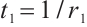 , then
, then 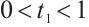 and
and 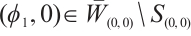 . Thus we have
. Thus we have 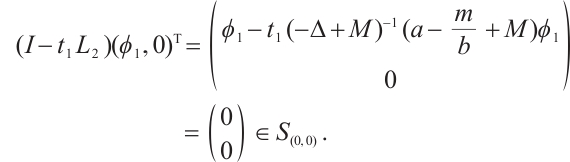
Therefore, 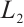 has
has 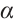 -property. By Lemma 4, we have
-property. By Lemma 4, we have 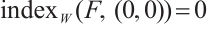 .
.
(iii) Obviously, (3) has a semi-trivial solution 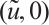 . Notice that
. Notice that 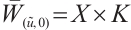 ,
, 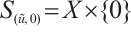 , we have
, we have 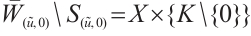 . Let
. Let 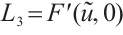 . There holds
. There holds
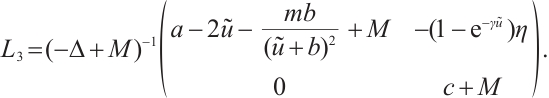
Assume that there exists 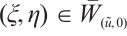 and
and 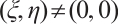 such that
such that 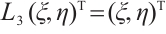 . Then
. Then
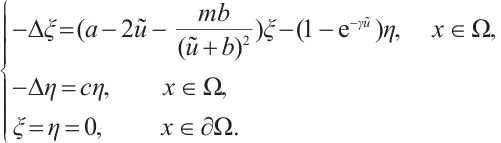
By 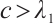 , we have
, we have 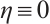 in
in 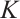 . If
. If 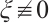 , then the above boundary value problem can be written as
, then the above boundary value problem can be written as
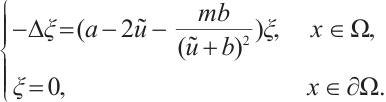
According to 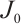 is invertible (see Lemma 5), we have
is invertible (see Lemma 5), we have  . This contradiction leads to
. This contradiction leads to 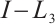 being invertible on
being invertible on 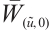 .
.
By Lemma 1 and 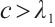 , we also have
, we also have
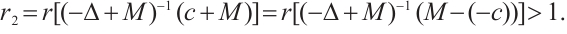
Notice that 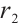 is the principal eigenvalue of
is the principal eigenvalue of 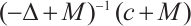 and the corresponding eigenfunction
and the corresponding eigenfunction 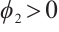 . Take
. Take 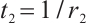 , then
, then 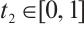 . Thus
. Thus 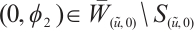 , and
, and
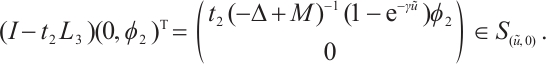
It shows  has
has 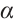 -properties. According to Lemma 4, there holds
-properties. According to Lemma 4, there holds 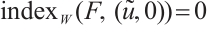 .
.
(iv) It follows from the proof of (iii) that 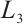 is invertible on
is invertible on 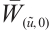 .
.
Now we claim 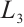 has no
has no 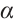 -properties on
-properties on 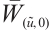 . If it is not true, then there exist
. If it is not true, then there exist 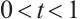 and
and 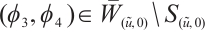 such that
such that 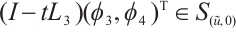 . Thus
. Thus
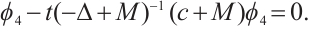
Notice that 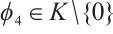 , then
, then 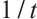 is one eigenvalue of the
is one eigenvalue of the 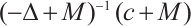 . On the other hand, by
. On the other hand, by 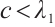 and Lemma 1 we have
and Lemma 1 we have 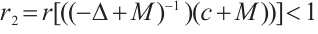 , which is a contradiction.
, which is a contradiction.
According to 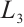 has no
has no 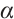 -property on
-property on 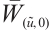 and Lemma 4, we have
and Lemma 4, we have
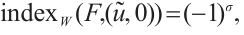
where 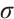 is the sum of the algebraic multiplicities of the eigenvalues of
is the sum of the algebraic multiplicities of the eigenvalues of 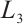 which are larger than one.
which are larger than one.
Assume that 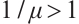 is the eigenvalue of
is the eigenvalue of 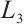 , and the corresponding eigenfunction is
, and the corresponding eigenfunction is 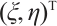 . Then
. Then 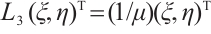 . This can be written as
. This can be written as
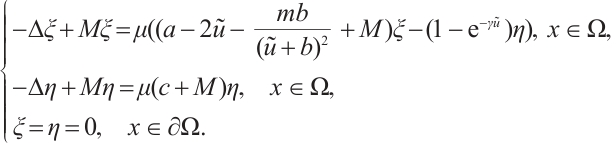
If 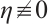 , then the inequality
, then the inequality
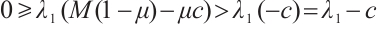
holds from the second equation of the above boundary value problem. This is a contradiction with 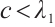 . Thus
. Thus 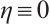 . So
. So 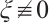 . Then from the first equation of the above boundary value problem we have
. Then from the first equation of the above boundary value problem we have
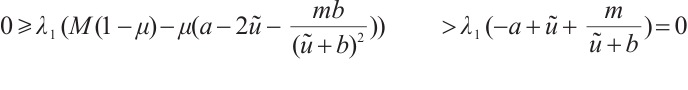
This is an obvious contradiction. Therefore,  has no eigenvalue greater than one, that is
has no eigenvalue greater than one, that is  and then
and then
It is similar to Lemma 6 that we can get Lemma 7.
Lemma 7 Let 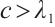 .
.
(i) If 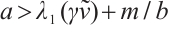 , then
, then 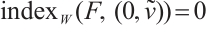 ,
,
(ii) If 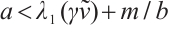 , then
, then 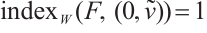 ,
,
where 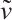 and
and 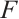 are given by (6) and (7), respectively.
are given by (6) and (7), respectively.
Theorem 3 If 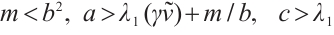 , then (3) has at least one positive solution.
, then (3) has at least one positive solution.
Proof From the additivity of the degree, combining Lemma 6 and Lemma 7 we have
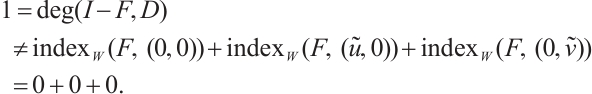
Therefore, (3) has at least one positive solution.
Remark 2 Theorem 3 shows that predator and prey can coexist as long as the Allee effect constant satisfies the appropriate conditions and the growth rates of predator and prey are appropriately large.
4 Uniqueness and Stability
In this section, we use the stability theory of linear operators to discuss uniqueness and stability of positive steady-state solutions. First, the following Lemma 8 is given.
Lemma 8 Let 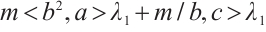 . There exists a constant
. There exists a constant 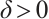 small enough, such that any positive solution of (3) is nondegenerate and linearly stable when
small enough, such that any positive solution of (3) is nondegenerate and linearly stable when 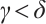 (if the positive solution exists).
(if the positive solution exists).
Proof Assume that it is not true. For 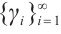 with
with  , there exists a sequence of positive solutions
, there exists a sequence of positive solutions 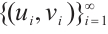 of (3), which are degenerate or unstable.
of (3), which are degenerate or unstable.
Now we suppose there are 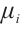 and
and 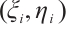 , which satisfy
, which satisfy 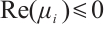 and
and 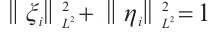 , such that
, such that
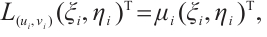
where 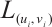 is the linearization operator of (3) at
is the linearization operator of (3) at 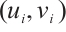 , i.e.,
, i.e.,
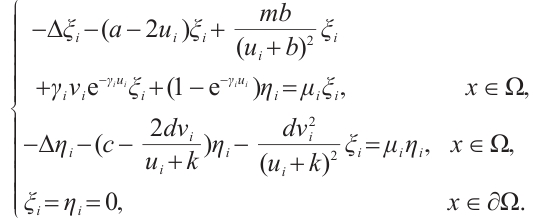 (8)
(8)
Obviously, 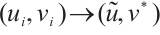 as
as  , where
, where  is a unique positive solution of the boundary value problem
is a unique positive solution of the boundary value problem
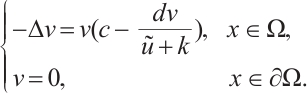
Multiplying 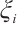 and
and 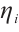 both sides of the first and second equations in (8), respectively, then integrating on
both sides of the first and second equations in (8), respectively, then integrating on 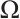 and adding the results, we can get
and adding the results, we can get
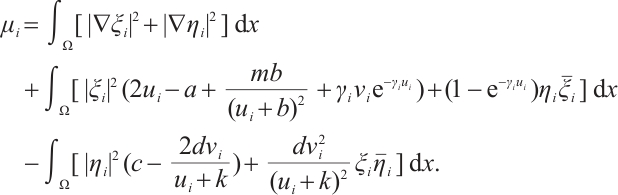
Note that 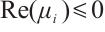 ,
, 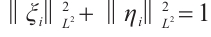 and
and 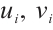 are both bounded according to Theorem 2. Thus
are both bounded according to Theorem 2. Thus 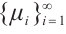 is also bounded. Suppose
is also bounded. Suppose 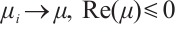 (take subsequences if necessary). By
(take subsequences if necessary). By 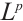 estimates for (8), both
estimates for (8), both 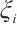 and
and 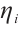 are also bounded in
are also bounded in 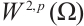 for
for 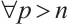 . So there exists a convergent subsequence of
. So there exists a convergent subsequence of 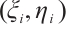 , which is still denoted by
, which is still denoted by 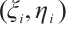 for the sake of convenience, such that
for the sake of convenience, such that 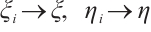 in
in 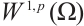 . Taking the limit in (8) with respect to
. Taking the limit in (8) with respect to  , then
, then 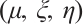 satisfies
satisfies
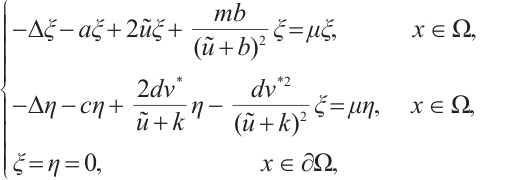 (9)
(9)
under the condition of weak convergence. According to the regularity theory, 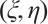 is a pair of classical solution of (9). It means that
is a pair of classical solution of (9). It means that 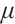 is a real number and
is a real number and 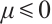 .
.
If 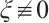 , then
, then 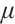 is an eigenvalue of the problem
is an eigenvalue of the problem
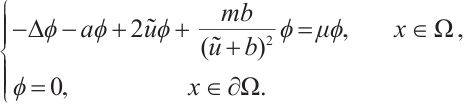
Combining with Lemma 5, there holds
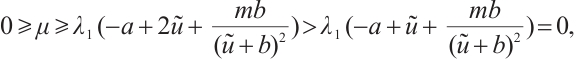
which is a contradiction. Thus  and (9) can be written as
and (9) can be written as
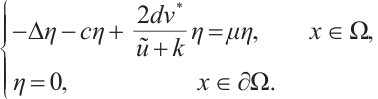
Similarly, if 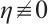 , then
, then
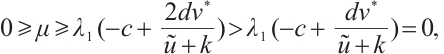
also a contradiction. So 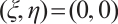 , which is a new contradiction with
, which is a new contradiction with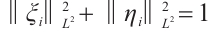 .
.
Theorem 4 Let 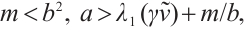
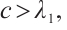 and
and 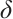 be a positive constant small enough, then (3) has a unique non-degenerate and linearly stable positive solution if
be a positive constant small enough, then (3) has a unique non-degenerate and linearly stable positive solution if 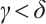 .
.
Proof By Theorem 3 and Lemma 8, the existence of positive solution is clear. So we only show the rest of this Theorem.
At first, it is easy to verify that both trivial solution 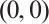 and semi-trivial solutions
and semi-trivial solutions 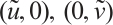 are all non-degenerate, linearly stable and isolated. According to the compactness theory[23], (3) has at most a finite number of positive solutions, which are recorded as
are all non-degenerate, linearly stable and isolated. According to the compactness theory[23], (3) has at most a finite number of positive solutions, which are recorded as 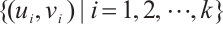 . It is similar to the proof of Lemma 8 that
. It is similar to the proof of Lemma 8 that 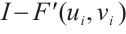 is invertible on
is invertible on 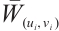 . Notice that
. Notice that 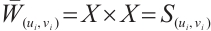 , we have
, we have 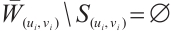 . Thus
. Thus 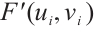 has no
has no 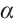 -property.
-property.
Furthermore, 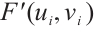 has no eigenvalue which is greater than 1. According to Lemma 4,
has no eigenvalue which is greater than 1. According to Lemma 4, 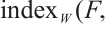
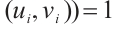 . From the additivity of degree and combining Lemma 6 and Lemma 7, we have
. From the additivity of degree and combining Lemma 6 and Lemma 7, we have
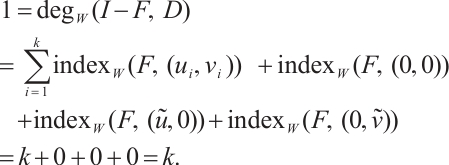
It follows that 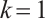 . The uniqueness of the positive solution is obtained.
. The uniqueness of the positive solution is obtained.
Remark 3 Theorem 4 shows that, as long as the Allee effect constant satisfies the appropriate relationship and the growth rate of predator and prey is appropriately large, the predator and prey not only coexist, but also the coexistence mode generated by low predator efficiency is stable.
5 Numerical Simulations
In this section, some numerical simulations for (2) and (3) in one-dimensional  will be carried out to verify the qualitative results of this paper. The algorithm used here is Pdepe in MATLAB. The initial value is taken as
will be carried out to verify the qualitative results of this paper. The algorithm used here is Pdepe in MATLAB. The initial value is taken as
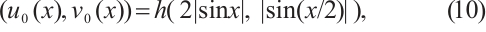 (10)
(10)
where 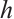 is a positive constant. The principal eigenvalue of
is a positive constant. The principal eigenvalue of 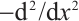 under the homogeneous Dirichlet boundary conditions is
under the homogeneous Dirichlet boundary conditions is 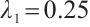 when
when 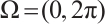 [24]. By the property of the principal eigenvalue we have
[24]. By the property of the principal eigenvalue we have
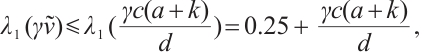
and
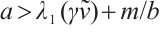 only if
only if 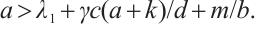
(i) Existence of steady-state solutions
As is well known, when the solution of (2) does not change with time, it is called the steady-state solution of (2), which is the solution of (3). The other remark here is that the weak Allee effect constant relationship 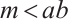 can be satisfied when
can be satisfied when 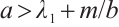 . A large number of numerical simulations are consistent with Theorem 3. Some examples are provided in the following statements, where the parameters are given by
. A large number of numerical simulations are consistent with Theorem 3. Some examples are provided in the following statements, where the parameters are given by
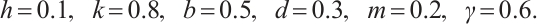
Let 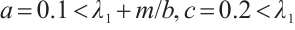 . (3) has a unique solution
. (3) has a unique solution  , which is shown in Fig. 1.
, which is shown in Fig. 1.
 |
Fig. 1 Steady-state solution 
|
Let 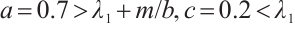 . (3) has a unique semi-trivial solution
. (3) has a unique semi-trivial solution  , which is shown in Fig. 2.
, which is shown in Fig. 2.
 |
Fig. 2 Steady-state solution 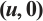
|
Let 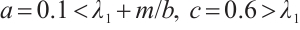 . (3) has a unique semi-trivial solution
. (3) has a unique semi-trivial solution 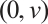 , which is shown in Fig. 3.
, which is shown in Fig. 3.
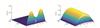 |
Fig. 3 Steady-state solution 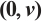
|
Let 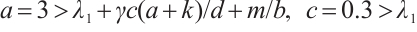 , (3) has a positive solution
, (3) has a positive solution 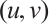 , which is shown in Fig. 4.
, which is shown in Fig. 4.
 |
Fig. 4 Steady-state solution 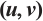
|
(ii) Influence of 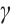 on the positive steady-state solutions
on the positive steady-state solutions
A large number of numerical simulations will verify the existence of steady-state solution when 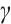 is sufficiently small. One of examples is shown in Fig. 5, which is consistent with the existence of positive solution in Theorem 4, where the parameters take
is sufficiently small. One of examples is shown in Fig. 5, which is consistent with the existence of positive solution in Theorem 4, where the parameters take
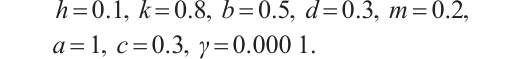 (11)
(11)
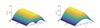 |
Fig. 5 Existence of steady-state solutions, 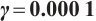
|
Furthermore, a large number of numerical simulations show that predator and prey density decrease with the increase of predation rate 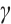 . One of the examples is shown in Fig. 6, where
. One of the examples is shown in Fig. 6, where 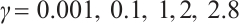 and the other parameters are given by (11).
and the other parameters are given by (11).
 |
Fig. 6 Existence of steady-state solutions, 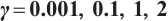 and and 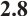
|
In addition, our numerical simulations show that under appropriate parameters, (3) has still the positive steady-state solution even if the 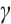 is large, which needs to be theoretically proved in future research. One example is shown in Fig. 7, where the parameters take
is large, which needs to be theoretically proved in future research. One example is shown in Fig. 7, where the parameters take
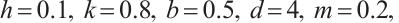
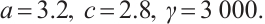 (12)
(12)
 |
Fig. 7 Existence of steady-state solutions, 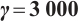
|
(iii) Influence of 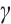 on the stability of positive steady-state solutions
on the stability of positive steady-state solutions
We change the initial value parameter 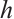 to simulate the disturbance of the initial value. For the convenience of discussion, the maximum norm of
to simulate the disturbance of the initial value. For the convenience of discussion, the maximum norm of 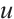 with respect to
with respect to 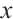 (denote it by
(denote it by 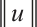 ) is plotted by Pdepe. If
) is plotted by Pdepe. If 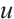 and
and 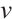 do not change with initial values after a long period of time, then
do not change with initial values after a long period of time, then 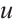 and
and 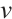 are stable. Otherwise, the steady-state solution is unstable. In fact, a large number of numerical simulations show that a positive steady-state solution is stable regardless of whether the
are stable. Otherwise, the steady-state solution is unstable. In fact, a large number of numerical simulations show that a positive steady-state solution is stable regardless of whether the 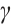 is large or small.
is large or small.
For example, let 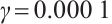 and
and 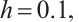
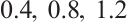 , the other parameters are given by (11), see Fig. 8. This shows that the positive solution is stable, which is consisted with the stability of positive solution in Theorem 4.
, the other parameters are given by (11), see Fig. 8. This shows that the positive solution is stable, which is consisted with the stability of positive solution in Theorem 4.
 |
Fig. 8 Stability of steady-state solutions, 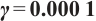
|
Let 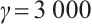 and
and 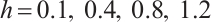 , the other parameters are given by (12), see Fig. 9. This shows that the positive solution is also stable, but the theoretical proof of this conclusion needs to be studied further.
, the other parameters are given by (12), see Fig. 9. This shows that the positive solution is also stable, but the theoretical proof of this conclusion needs to be studied further.
 |
Fig. 9 Stability of steady-state solutions, 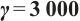
|
6 Conclusion
We simulate the interaction between invertebrates and plankton using a modified Leslie-Gower predator-prey model with the Ivlev type functional response function, which is used to describe the fact that Tortanus dextrilubatus prey on zooplankton. Meanwhile, we also focus on the Allee effect in the model. Our research shows that under the Allee effect, predators and prey can coexist. Specifically, the growth rates of predators and prey can control the uniqueness of the coexistence pattern, which is consistent with the actual situation. We employ some numerical simulations primarily to verify the rationality of the conditions in this article, such as Lemma 8 and Theorem 4, etc. At the same time, we also verify the stability of the steady-state solutions by making small perturbations to the initial values. In a sense, these studies only provide a framework for model dynamics, and there are still some things that have not been studied, such as simulating the non-enclosure of habitats using Neumann boundary conditions, then the properties of constant and non-constant solutions of the model will be an interesting problem. In fact, more complex coexistence patterns of predator-prey systems can be examined through Turing pattern and Hopf bifurcation.
References
- Zhang C H, Yang W B. Dynamic behaviors of a predator-prey model with weak additive Allee effect on prey[J]. Nonlinear Analysis: Real World Applications, 2020, 55: 103137. [Google Scholar]
- Aguirre P, González-Olivares E, Sáez E. Three limit cycles in a Leslie-Gower predator-prey model with additive Allee effect[J]. SIAM Journal on Applied Mathematics, 2009, 69(5): 1244-1262. [Google Scholar]
- Cai Y L, Banerjee M, Kang Y, et al. Spatiotemporal complexity in a predator: Prey model with weak Allee effects[J]. Mathematical Biosciences and Engineering, 2014, 11(6): 1247-1274. [Google Scholar]
- Pal P J, Mandal P K. Bifurcation analysis of a modified Leslie-Gower predator-prey model with Beddington-DeAngelis functional response and strong Allee effect[J]. Mathematics and Computers in Simulation, 2014, 97: 123-146. [Google Scholar]
- Yang W S, Li Y Q. Dynamics of a diffusive predator-prey model with modified Leslie-Gower and Holling-type Ⅲ schemes[J]. Computers & Mathematics with Applications, 2013, 65(11): 1727-1737. [Google Scholar]
- Yang W B. Analysis on existence of bifurcation solutions for a predator-prey model with herd behavior[J]. Applied Mathematical Modelling, 2018, 53: 433-446. [Google Scholar]
- Li S B, Wu J H, Dong Y Y. Uniqueness and stability of a predator-prey model with C-M functional response[J]. Computers & Mathematics with Applications, 2015, 69(10): 1080-1095. [Google Scholar]
- Hooff R C, Bollens S M. Functional response and potential predatory impact of Tortanus dextrilobatus, a carnivorous copepod recently introduced to the San Francisco Estuary[J]. Marine Ecology Progress Series, 2004, 277: 167-179. [Google Scholar]
- Wang X C, Wei J J. Dynamics in a diffusive predator-prey system with strong Allee effect and Ivlev-type functional response[J]. Journal of Mathematical Analysis and Applications, 2015, 422(2): 1447-1462. [Google Scholar]
- Wang H L, Wang W M. The dynamical complexity of a Ivlev-type prey-predator system with impulsive effect[J]. Chaos, Solitons & Fractals, 2008, 38(4): 1168-1176. [Google Scholar]
- Wang X C, Wei J J. Diffusion-driven stability and bifurcation in a predator-prey system with Ivlev-type functional response[J]. Applicable Analysis, 2013, 92(4): 752-775. [Google Scholar]
- Li S B, Wu J H, Nie H. Steady-state bifurcation and Hopf bifurcation for a diffusive Leslie-Gower predator-prey model[J]. Computers & Mathematics with Applications, 2015, 70(12): 3043-3056. [Google Scholar]
- Zhou J, Shi J P. The existence, bifurcation and stability of positive stationary solutions of a diffusive Leslie-Gower predator–prey model with Holling-type Ⅱ functional responses[J]. Journal of Mathematical Analysis and Applications, 2013, 405(2): 618-630. [Google Scholar]
- Cai Y L, Zhao C D, Wang W M, et al. Dynamics of a Leslie-Gower predator-prey model with additive Allee effect[J]. Applied Mathematical Modelling, 2015, 39(7): 2092-2106. [Google Scholar]
- Yang L, Zhong S M. Dynamics of a diffusive predator-prey model with modified Leslie-Gower schemes and additive Allee effect[J]. Computational and Applied Mathematics, 2015, 34(2): 671-690. [Google Scholar]
- Ma Z P. Spatiotemporal dynamics of a diffusive Leslie-Gower prey-predator model with strong Allee effect[J]. Nonlinear Analysis: Real World Applications, 2019, 50: 651-674. [Google Scholar]
- Wang L J, Jiang H L. Properties and numerical simulations of positive solutions for a variable-territory model[J]. Applied Mathematics and Computation, 2014, 236: 647-662. [Google Scholar]
- Cano-Casanova S. Existence and structure of the set of positive solutions of a general class of sublinear elliptic non-classical mixed boundary value problems[J]. Nonlinear Analysis: Theory, Methods & Applications, 2002, 49(3): 361-430. [Google Scholar]
- Blat J, Brown K J. Global bifurcation of positive solutions in some systems of elliptic equations[J]. SIAM Journal on Mathematical Analysis, 1986, 17(6): 1339-1353. [Google Scholar]
- Ye Q X, Li Z Y, Wang M X, et al. Introduction to Reaction-Diffusion Equation[M]. Beijing: Science Press, 2013(Ch). [Google Scholar]
- Pao C V. Nonlinear Parabolic and Elliptic Equations[M]. Cham: Springer-Verlag, 1992. [Google Scholar]
- Wang M X. Nonlinear Elliptic Equation[M]. Beijing: Science Press, 2007(Ch). [Google Scholar]
- Du Y H, Lou Y. Some uniqueness and exact multiplicity results for a predator-prey model[J]. Transactions of the American Mathematical Society, 1997, 349(6): 2443-2475. [Google Scholar]
- Wang L J, Jiang H L. The multiplicity and uniqueness of positive solutions for a competition model with diffusion[J]. Journal of Wuhan University (Natural Science Edition), 2015, 61(4): 308-314(Ch). [Google Scholar]
All Figures
|
Fig. 1 Steady-state solution
|
| In the text | |
|
Fig. 2 Steady-state solution
|
| In the text | |
|
Fig. 3 Steady-state solution
|
| In the text | |
|
Fig. 4 Steady-state solution
|
| In the text | |
|
Fig. 5 Existence of steady-state solutions,
|
| In the text | |
|
Fig. 6 Existence of steady-state solutions, and
|
| In the text | |
|
Fig. 7 Existence of steady-state solutions,
|
| In the text | |
|
Fig. 8 Stability of steady-state solutions,
|
| In the text | |
|
Fig. 9 Stability of steady-state solutions,
|
| In the text | |
Current usage metrics show cumulative count of Article Views (full-text article views including HTML views, PDF and ePub downloads, according to the available data) and Abstracts Views on Vision4Press platform.
Data correspond to usage on the plateform after 2015. The current usage metrics is available 48-96 hours after online publication and is updated daily on week days.
Initial download of the metrics may take a while.